I am a short man. I have an even shorter girlfriend. Odds are, any offspring would be short. Odds are, their partners would be similar or smaller in height, especially if they are male. Does this mean I am destined to be the heir to a slowly shrinking troglodyte race, my pristine DNA squirted from troll to troll until it degrades to a corrosive, fetid broth? Let’s put the thesaurus away and have a look.
The model
On average, men are taller than women, though the heights of both populations approximately follow a Gaussian distribution

where 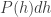

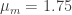

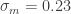

where I’ve used 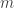


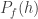

For the sake of simplicity, let’s consider a simple case. A single man and a single woman meet, producing a single male and single female. The parents are then deleted from the population, so the time evolution of the population depends on how the heights of the parents 
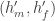

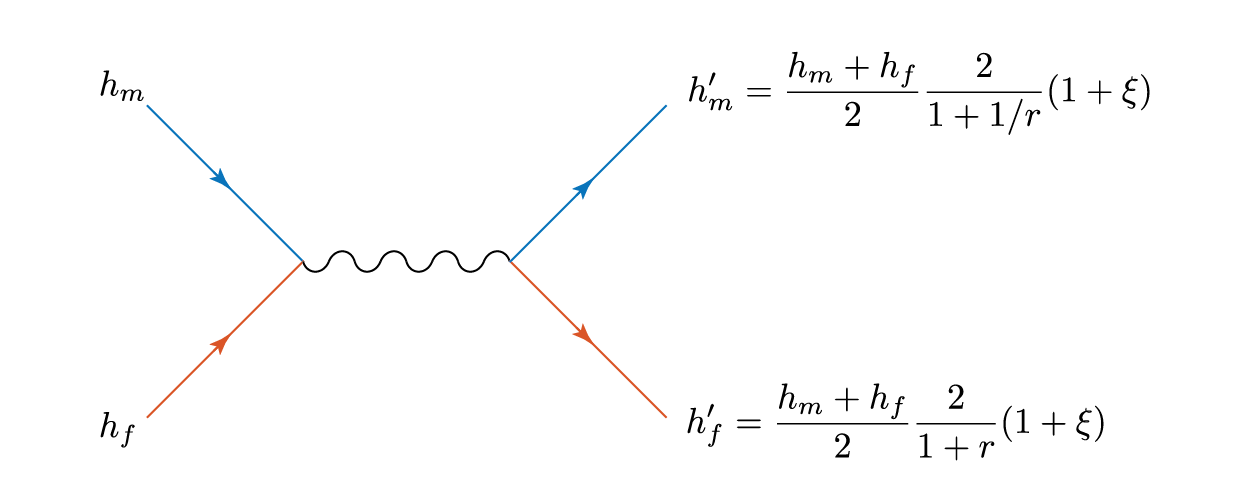
then both children are the same height – the average of the two parents. Let’s physics this up a bit, as we have two things interacting and effectively ‘scattering’ away with different properties. We can represent this as a Feynmann diagram, where blue represents men and orange women (yes, even the default Matlab plot colours suffer from tired gender stereotypes).
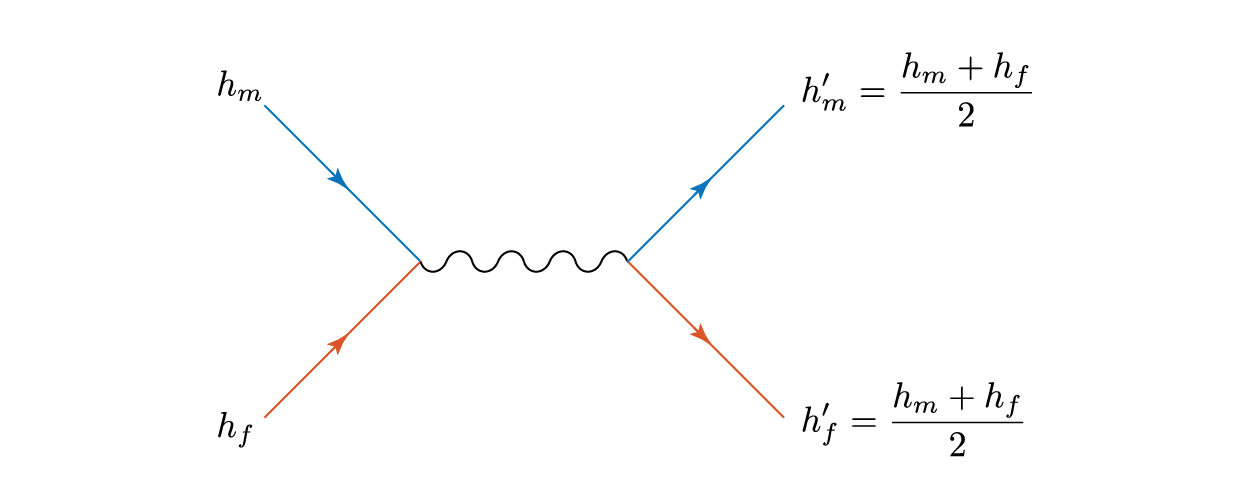
The squiggly line represents the, er, interaction, and the childish amongst you can reinterpret gluons or propagators as you wish.
Total height is conserved in this process, and there is an effective transfer of height

from the man to the woman.
Equal height children
What effect does this have on the population? I simulate a large number of these interactions, randomly picking parents and adjusting their properties as above:
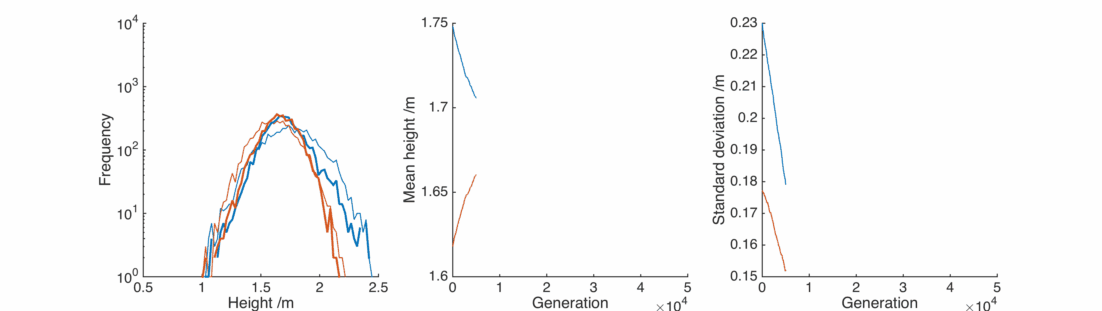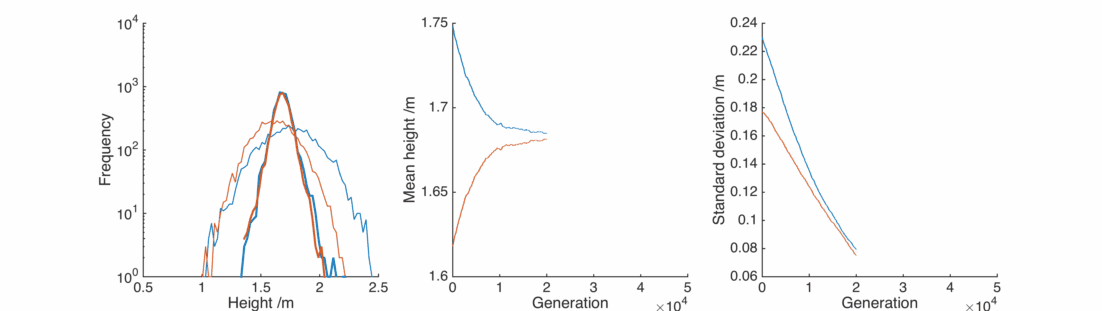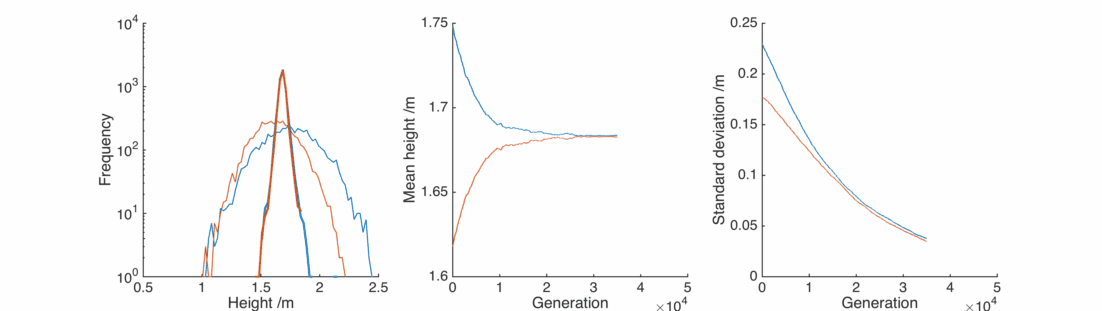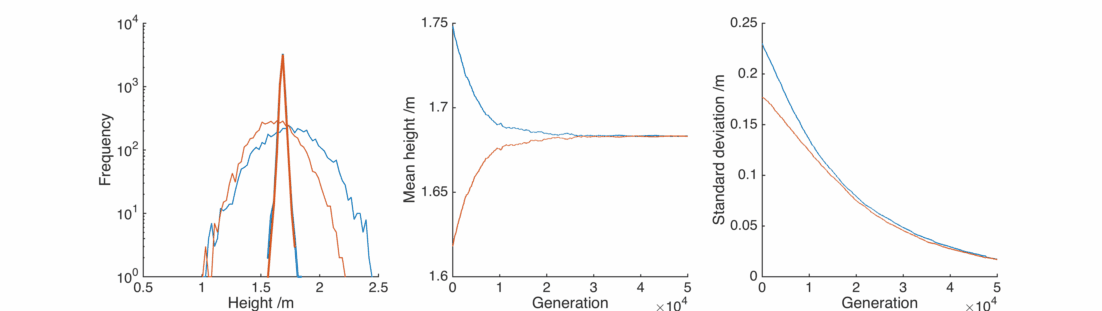
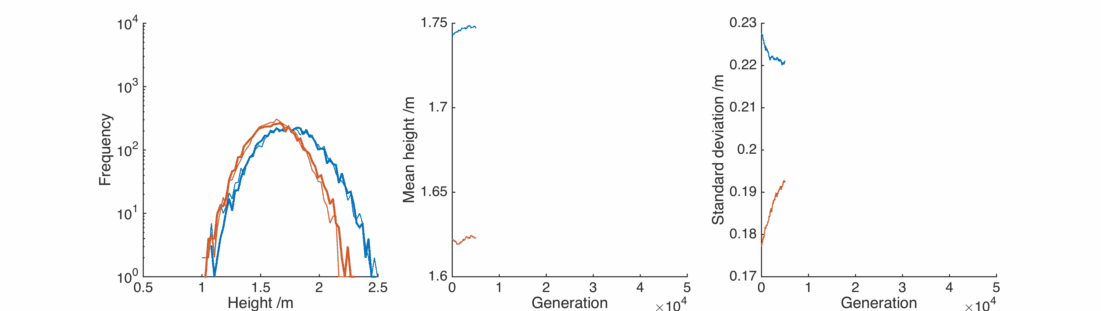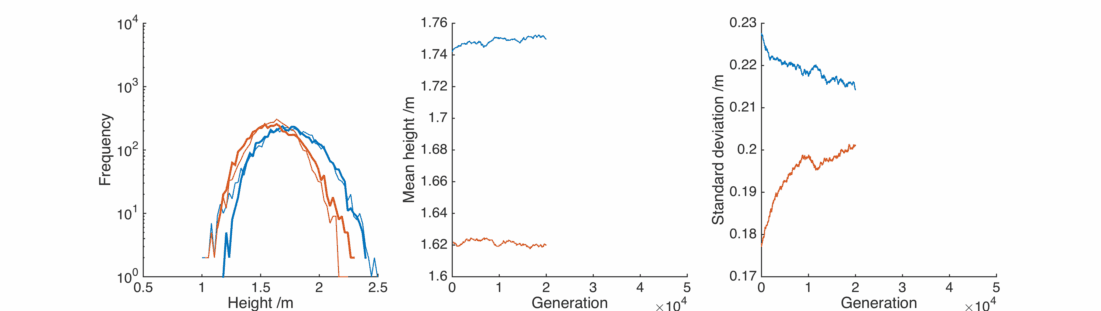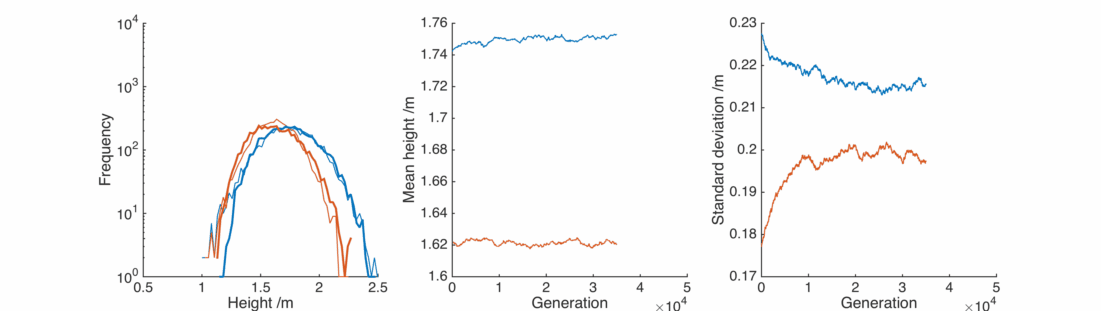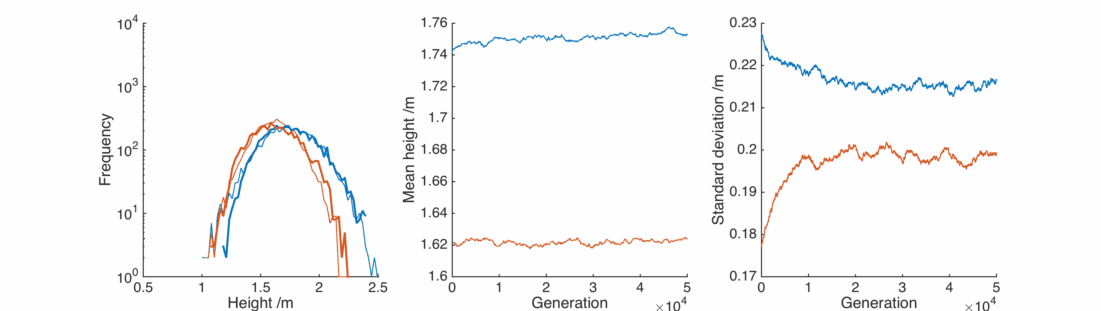
This doesn’t look good – very rapidly the male and female populations converge to the same average height, and the variance of heights within each population is dropping towards zero. This isn’t surprising – if you replace two quantities by their average, the distribution will be losing people at the edges and gaining them in the middle.
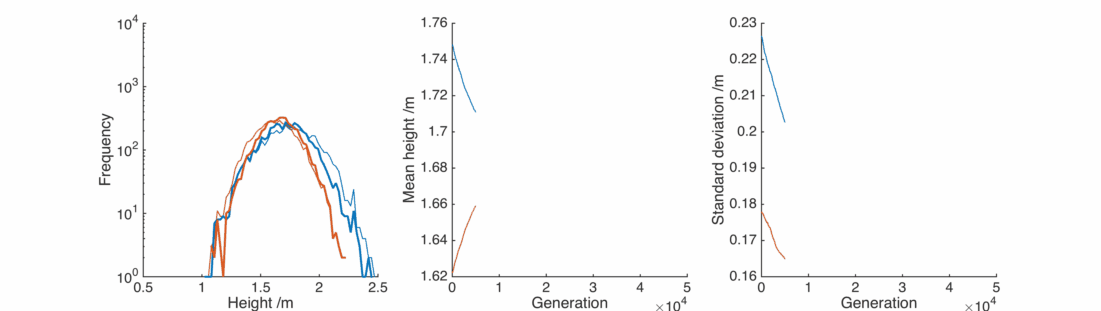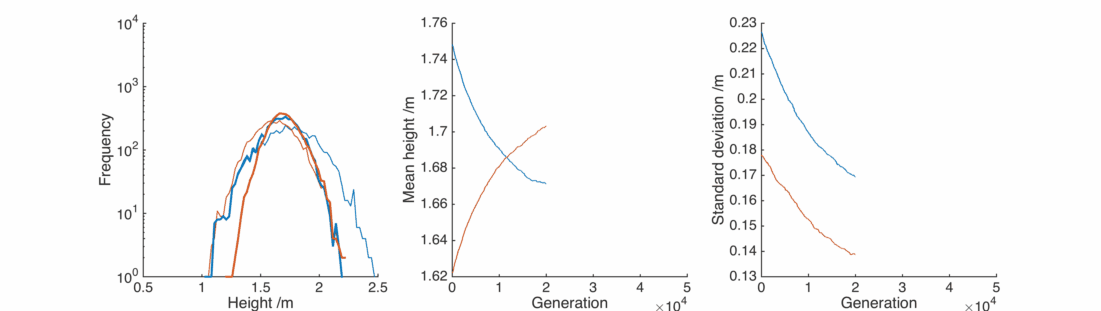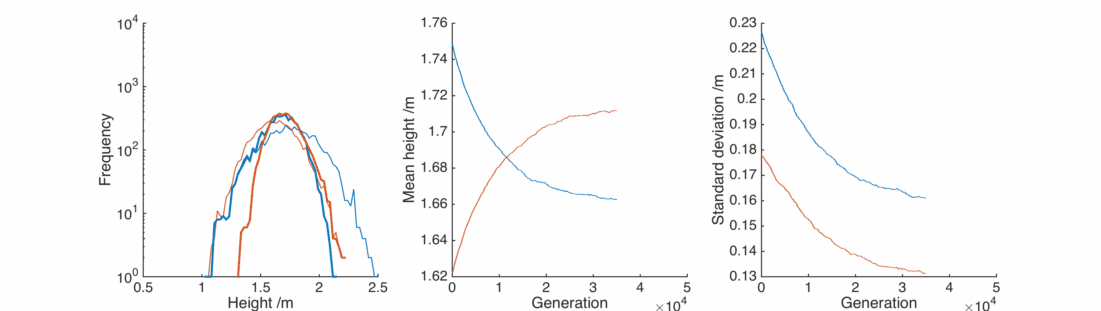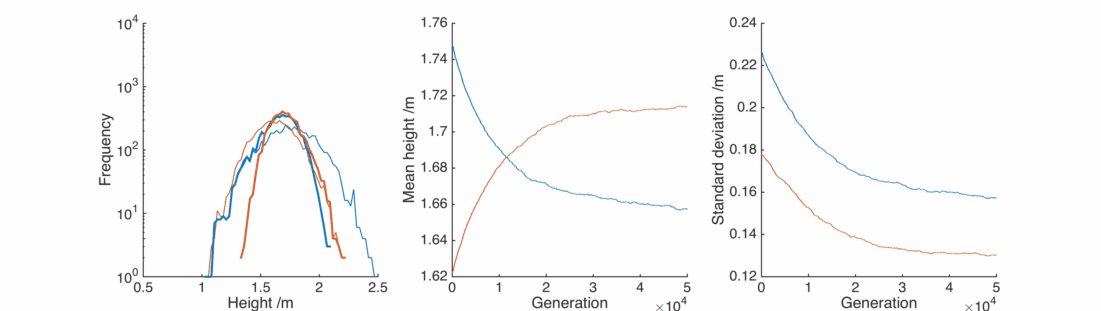
Asymmetric children
Let’s adjust our scattering procedure with the assumption that the male child will tend to be taller than the female child, as we observe in the real population:

To see that this is appropriate, consider the average height of the male child averaged over the population (where 

which by definition of the mean heights

So if we take that 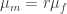

and the average male child height should follow the same ratio on average. What does this do to the population evolution then?
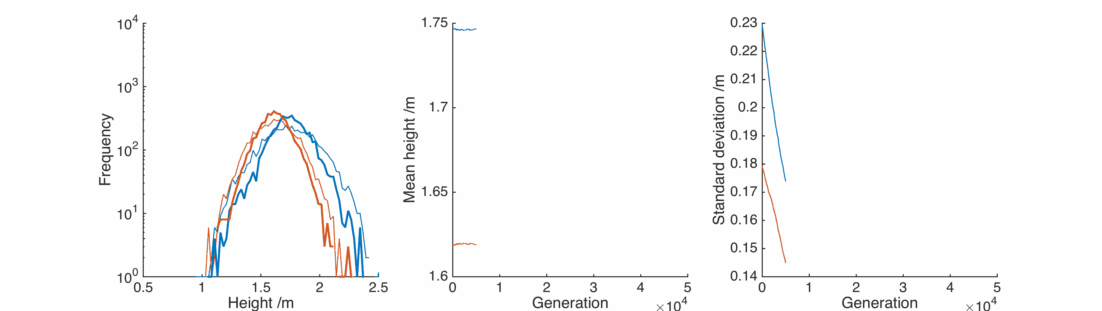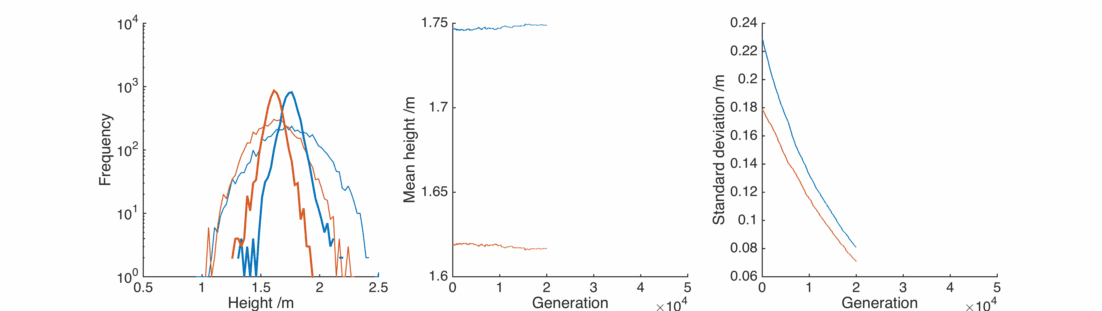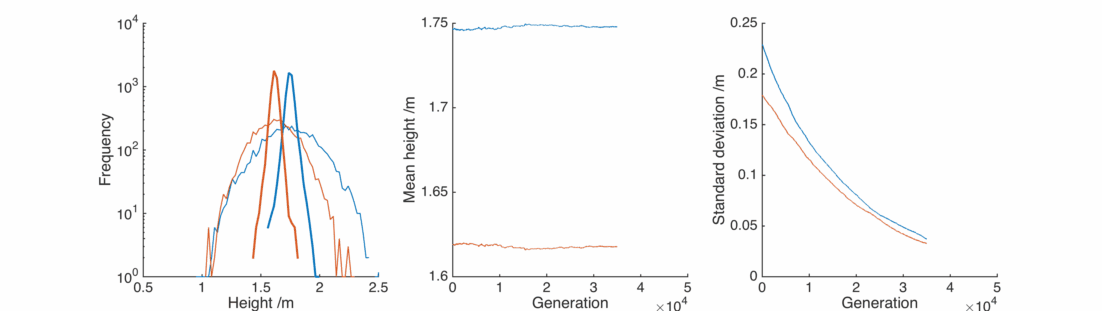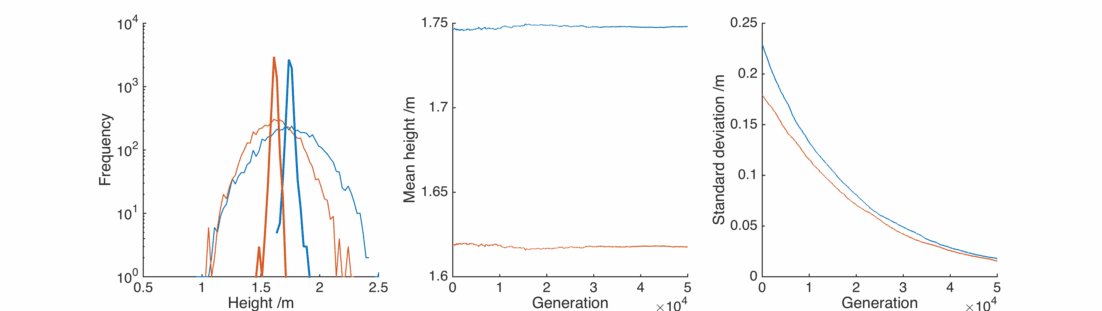
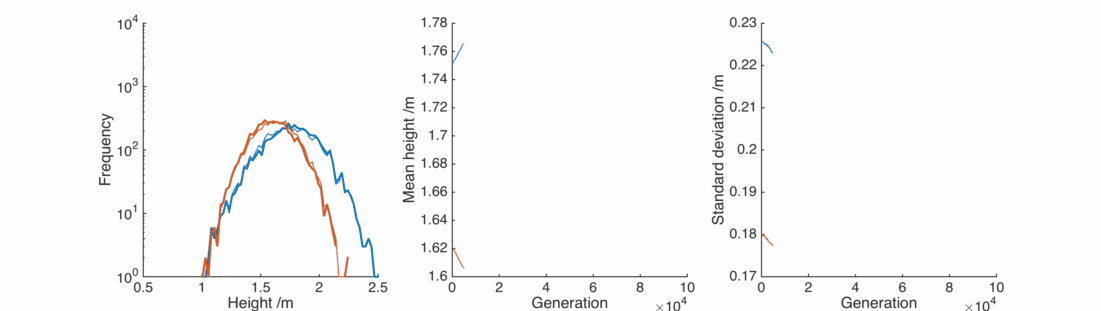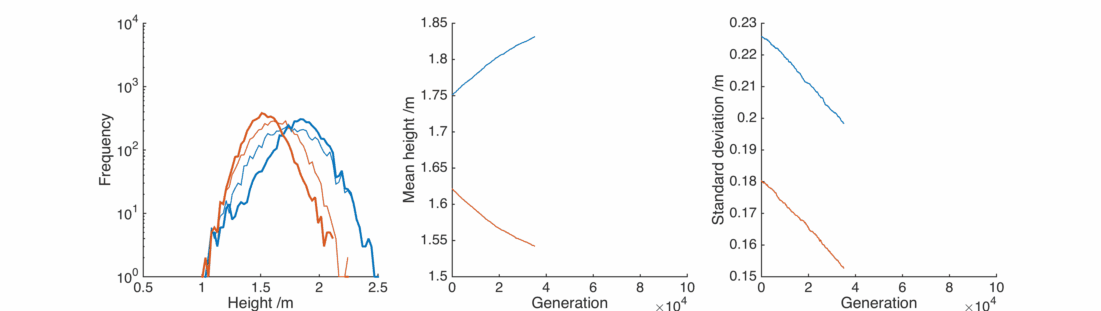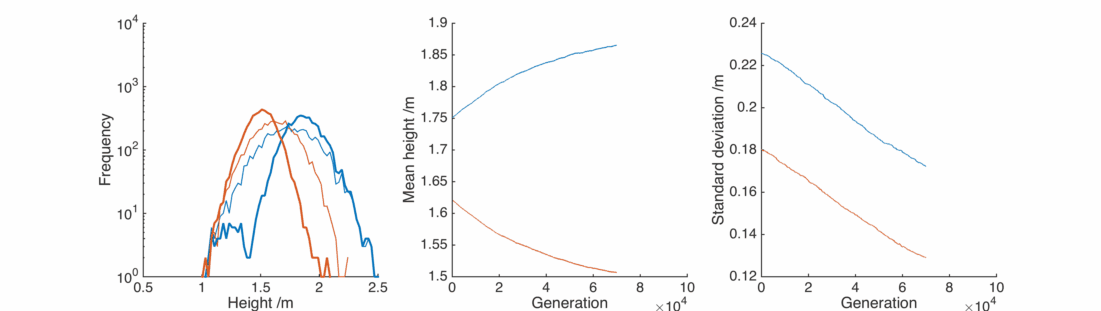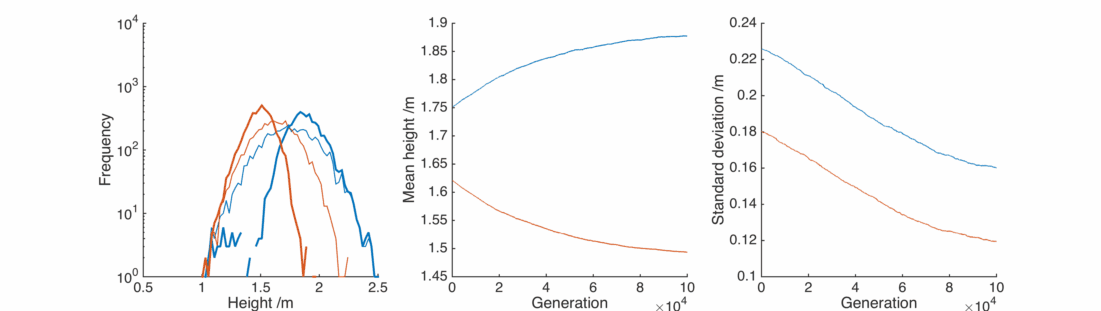
Great, the average heights stay pretty much exactly the same as we wanted. By adjusting the 

However, the populations still don’t resemble what we observe in the real world. The repeated averaging process homogenises the populations in an unrealistic way.
Random children
What we need is a bit of randomness – adjust the scattering procedure so that it reads
where 

This is nice, with a bit of gender bias in child heights and a sprinkling of randomness we can reproduce the observed population, and it’s relatively stable.
Interaction probabilities
To start answering the question I posed at the beginning, we’ll need to adjust the final part of the scattering process – the likelihood of interaction 
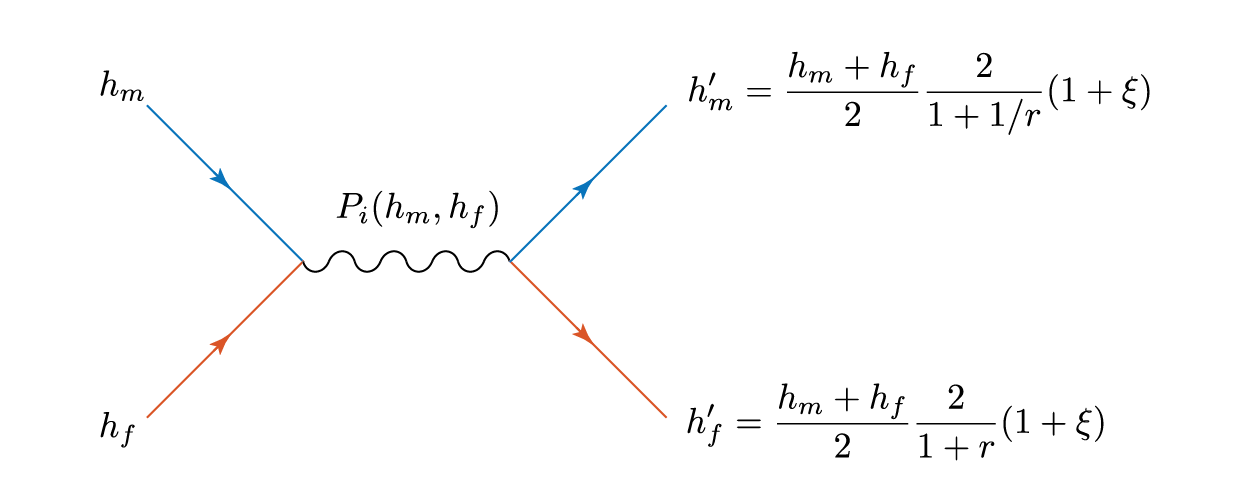
This is, in general, a function of the heights of the two prospective parents. It could take a number of forms. For example, what if a couple couldn’t tolerate a height difference of more than, say, 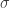

would encourage pairings where the heights of the parents are similar, and discourage pairings with a large different in height. What does this look like in practise?
Here the real winners are short men and tall women, whose populations significantly overlap. They breed themselves out of existence, leaving a lonely population of tall men and short women, each repulsed by the other and breeding slows down.
This is unrealistic, what about the situation where women will tend towards taller men? Here, set 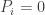

In this case the situation is reversed, somewhat counter-intuitively the female population gets taller as it preferentially interacts with taller men. The male population gets shorter as the taller section breeds itself away, In general everyone is pretty happy, except for the initial male population below 1.25 m or so.
PDE model
The above procedure is pretty simple to conceptualise and implement. However, it can be quite slow, especially when the aggregate probabilities of interaction are low. There is another way to look at this process, motivated by physics (naturally).
It is often natural to consider the evolution of a distribution function of some species under the influence of internal and external forces. In plasma physics, for example, the distribution function is that of a particle species over position and velocity. The Vlasov equation then dictates how this distribution function evolves. An important component is the description of scattering processes – when two particles collide they change their velocities, and therefore the shape of the velocity distribution function is changed. One can show that the distribution function is unchanged by scattering (on average) when it attains a Maxwell-Boltzmann form, i.e. is in thermodynamic equilibrium.
It is interesting to consider how our two distribution functions interact through scattering, and under what conditions the populations might be stable. I’ll just think about the simple case here when 
The value of 

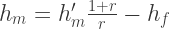
In the range 


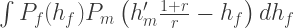
The rate at which
Overall then, we have
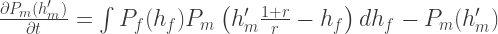
and analogously for the female population
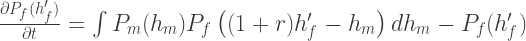
By solving these integro-differential equations numerically, we get the same behaviour as was seem earlier, namely narrowing of the two populations around their respective means.
Hand-waving alert
Notice that the equations above take the form of convolutions. If we Fourier transform with respect to the primed variables then, we have


(neglecting some constants and lots of rigour). A non-trivial stable solution then arises when the Fourier transform of the two distributions is 1, or when the distributions themselves tend to a delta-function. This is the limiting behaviour we observe in numerical simulation.
I’m not comfortable enough with this technique to take it much farther though, and would therefore welcome corrections or pointers to useful resources.

Hi there! I love your blog entries – I’ve been following for about a year. I love how you start with a story, and build up the mathematics to reflect some new parameter that is added to make things more realistic.
LikeLike
Thanks! The usual procedure is to idly fiddle around with some maths until I get something interesting. Of course I only write about the ones that turn out tractable, so there is some publication bias too 🙂
LikeLike
Oh man, I love the Feynmanesque diagramming. This is a fantastic post!
LikeLike
Haha, thanks! You have to be a special kind of geeky to start at reproduction and end at Feynman (ish) diagrams.
LikeLike
In the simplest case, how does the height the female population not equal the height of the male population after one generation? In other words, in the case where both offspring have the same height, after one generation the male and female population will have the same samples, however your figures do not show this.
If I am not mistaken. To do this you first sample the initial population. Randomly permute the samples to choose the parents. Then compute the average height to get the heights of the new females and males, and then repeat the procedure.
LikeLike
Hi, the reason is that the initial populations contain a broad spread of heights. The two children are the same height for a given set of parents, but over the distribution of parents there will still be many children of different heights after a generation. In each ‘generation’ I pick a sample of parents at random, and replace them with their averages. Doing this enough times the variance in the population is reduced, and I show at the end of the post how the limit of this process is a delta function in height.
LikeLike